





To the Editors:
Asthma is a complex disease characterised by inflammation and remodelling of the airways. Over the past few decades enormous progress has been made to understand which genes are associated with asthma development and several interactions between genes and environmental factors have been elucidated. Investigations into genetic and gene expression profiling, as well as single nucleotide polymorphism analyses, have helped to better understand the underlying molecular mechanisms of asthma. However, the recent identification of novel regulatory functions for transposable and transposed genetic elements (TEs) may be an important and new key to help understand the genetics that cause the heterogeneous manifestations of the asthma pathology.
Approximately 45% of the human genome is made of TEs [1]. The vast majority of TEs originate from retrotransposition of genetic elements known as short and long interspersed nuclear elements, long terminal repeat-superfamilies and direct transposition of TE-containing genomic DNA. Formerly regarded as junk DNA, it is now becoming increasingly evident that TEs often function to regulate and fine tune gene expression [2, 3]. TE-driven transcription frequently controls the expression of protein coding genes via alternative promoters, cis-regulatory non-protein-coding RNAs and through the formation of double stranded short RNAs. Faulkner and Carninci [3] demonstrated that transcription initiation from promoters present in TEs is a general phenomenon, even when they are corrupted and not easily recognised as genuine transposons. In addition, TE insertions into gene sequences affect RNA stability and splicing variants [4, 5]. Approximately 30% of human mRNA contains at least one retrotransposon, and the mRNA levels were shown to be inversely proportional to the percentage of retrotransposons [6]. Such findings suggest that TEs are intrinsic components of the transcriptional region of protein coding loci with crucial regulatory effects. Furthermore, these novel data imply a control mechanism of unknown character that selectively and specifically contributes to a cell-type specific accumulation of TEs. At the level of the gene structure, TEs reshape the genes by 1) insertion mutagenesis; 2) repair of DNA double stranded breaks; and recombination between non-allelic homologous elements by 3) deletions; 4) duplications; and 5) inversions. TE-encoded proteins often have DNA-binding or modifying functions, such as copying, breaking, joining, and/or degradation of the double helix. Transposed in the wrong position, however, TEs may cause the development of (complex) diseases. Specific expression patterns of TEs in germ-line and cancer cells have been associated with numerous human diseases, usually due to novel insertions across protein coding exons [7].
Asthma is a very heterogeneous disease. Often asthma is associated with atopy, but not all atopic individuals develop asthma. Some asthma patients outgrow symptoms, whereas others acquire asthma later in life. Still other patients develop the disease in early childhood and suffer from asthma their entire life. In addition, ethnicity, sex and age affect asthma susceptibility and severity [8, 9]. How can we explain these different patterns? A challenging hypothesis may be that the heterogeneity of asthma is due to differences with respect to TE-regulated asthma-susceptibility genes. TEs are not only inherited, but they may also be acquired during a lifetime. The number of repetitive TE sequences varies between different human populations and between individuals. Duplications and deletions, as well as inversions, are facilitated by recombination between homologous regions of TE sequences in different parts of the genome. The highly dynamic nature of TEs is demonstrated by retrotransposon copy-number variations (representing deletions and/or insertions) between human populations and even among individuals [2]. The dynamic nature of TEs, and the observation that they function as mobile genetic switches, make them ideal candidates to explain the heterogeneity and complexity of the asthma phenotype. TE dynamics may explain why the prevalence of asthma has rapidly increased over the past few decades. TEs may also explain why people can acquire the disease during a lifetime. Because of the copy–paste mechanism, newly acquired TEs will afterwards become a heritable part of the genome. It remains to be established, however, whether the dynamics of TEs can be modulated by environmental cues or whether it behaves purely stochastically.
Thus far, 12 asthma susceptibility genes have been identified by either positional cloning (DPP10, CYFP2, HLAG, GPRA, SFRS8, PHF11, ADAM33, PCDH1) or genome-wide association (CH13L1, ORMDL3, PDE4D, DENND1B) [10]. These genes may comprise the preferred starting point of investigations into TE-regulated asthma susceptibility genes. The expression of the 12 asthma susceptibility genes has been shown to be tissue-specific [10]. How tissue-specific gene expression is determined still remains a matter of intense scrutiny, but a remarkable coincidence is the tissue-specific regulatory action of TEs. Therefore, gene expression arrays may be an appropriate screening tool to find tissue-specific asthma genes.
Repetitive DNA elements may indeed affect the development of asthma in children. Studies have shown that genetic instability in the micro-satellite DNA is present in adult asthmatics, but also in asthmatic children [11, 12]. Historically, interleukin (IL)-3, IL-5 and granulocyte-macrophage colony-stimulating factor (GM-CSF) have been implicated in the development of asthma, in particular, because the expression of these genes has been shown to be up-regulated in several cell types of asthma patients and they are thought to cause eosinophilia. It is intriguing that IL-3, IL-5 and GM-CSF, which are found collinear on chromosome 5q31, are all regulated by the same upstream, yet unidentified, enhancer [13], but this finding is compatible with the novel functions attributed to TEs [6]. The presence of TEs within or in the vicinity of asthma susceptible genes could potentially function as transcription modulators of these genes. Therefore, it is important to precisely determine the position and number of TEs present in and around asthma susceptible genes, in both healthy controls and asthmatic patients, and evaluate the functional consequences of these genetic elements with respect to the transcriptome.
The final step in the detection of the TE-regulated transcriptome in healthy and diseased tissue is to assess whether the detected sequences have a biological function. Here, the main question is whether or not TEs within or in the vicinity of asthma susceptible genes modulate the gene expression of these genes. This can be performed using cap-analysis gene expression, a method that specifically and unequivocally identifies the exact transcription start sites of genes [6]. Once the regulated retrotransposon transcriptome of healthy tissue is known, it can be compared with that of asthma patients or additional complex diseases. This will provide unprecedented details about genes and gene expression patterns, and how they relate to the development and/or establishment of complex diseases. The impact of TEs on phenotype (variation) and disease are depicted in figure 1.
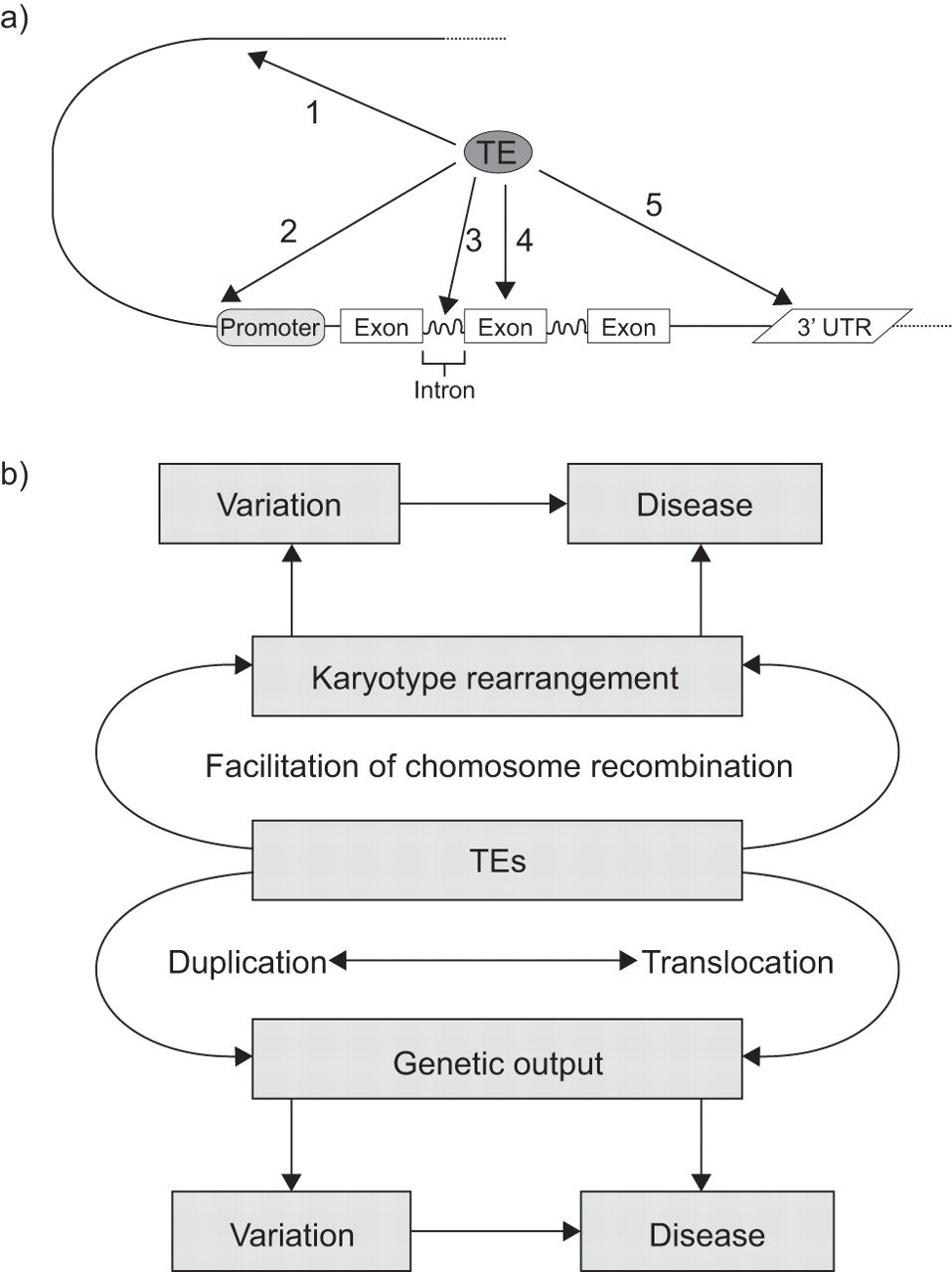
{kind=link}
a) Impact of transposable and transposed elements (TEs) on human genome structure and how they influence gene expression. TEs determine and fine tune genetic output and stability of mRNA due to their function as: 1) an enhancer; or 2) an alternative promoter; or by integration: 3) into introns; 4) into exons; and 5) into the untranslated regions (UTR). b) The role of TEs in the development of (complex) diseases. Due to amplification and/or translocation, TEs may jump within or into the vicinity of disease susceptible genes and start to interfere with normal genetic output. However, TEs may facilitate genetic rearrangements resulting in karyotype changes leading to genomic variation and/or diseases.
With the advent of next generation sequencing and rapid genome and transcriptome analyses, the dynamic nature of TEs and their role in shaping intra- and inter-individual variation may soon be resolved. Because these developments will dissect TE-induced variation at ever increasing resolution, they will lead to a better understanding of TEs and their overall impact on gene expression, human health and the (epi)genetic constitution that underlies complex lung diseases, including asthma. We have already identified the number, position and type of TEs present in the 12 aforementioned asthma-associated genes and we are in the process of screening the same panel of genes in asthma patients. It may well be that some phenotypes of asthma are predominantly associated with TE-regulated genes of the immunological compartment, whereas other phenotypes of asthma are linked to a specific set of TE-regulated genes in lung tissue cells. Therefore, cell- and phenotype-specific understanding of the healthy and diseased transcriptomes may lead to better treatment and to personalised medicine. We would like to encourage pulmonary scientists to further examine the almost entirely unexplored TE field, which integrates clinics, the laboratory and bioinformatics. The development of entire novel strategies to treat complex lung diseases should be envisaged.
Footnotes
Support Statement
M. Roth, M. Tamm and P. Borger received research grants from the Swiss National Science Foundation.
Statement of Interest
None declared.
- ©ERS 2012
REFERENCES










